Almost all eukaryotic protein coding genomes contain non-coding intervening sequences called introns. Spliceosomes are complex ribonuclear machines in eukaryotes that remove the non-coding introns from primary transcript, precursor mRNA (pre-mRNA or hnRNA).
Alternatively, the term spliceome can be used to describe the complete set of all possible alternative splices in an organism, analogous to the genome or proteome.
Spliceosomes are variably composed of as many as 300 distinct proteins and five RNAs, making them among the most complex macromolecular machines known. Free Full Text Article, review article abstract.
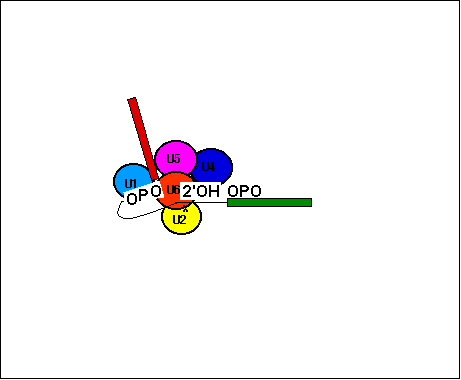
Essential components of the spliceosome are the small RNA-protein complexes called small nuclear ribonucleoproteins (snRNPs, pronounced 'snurps'). These are U1, U2, U4, U5, and U6, so named because they are rich in uridine nucleotides. In addition to snRNPs, splicing requires many non-snRNP protein factors. The snRNPs participate in several RNA-RNA and RNA-protein interactions.
The spliceosome recognizes specific conserved (5', 3', branch) splice sites on the pre-mRNA. The intronal area between these locations is excised, and the two exons are spliced (ligated).
Modified: "The spliceosome is a macromolecular machine that carries out the excision of introns from eukaryotic pre-mRNAs and splicing together of exons. Four large RNA–protein complexes, called the U1, U2, U4/U6 and U5 small nuclear ribonucleoprotein particles (snRNPs), and some non-snRNP proteins assemble around three short conserved (above) sequences within the intron in an ordered manner to form the active spliceosome.
The U1 and U2 snRNPs bind to the 5´ splice site and the branch point of pre-mRNA respectively, and a pre-assembled U4/U6·U5 tri-snRNP then joins these snRNPs to form the spliceosome [5–8,12]. The 5´ end of U1 snRNA is complementary to the conserved sequence at the 5´ splice site, and hence the recognition of the 5´ splice site is achieved partly through RNA base-pairing. The conserved sequence at the branch point also pairs with a complementary sequence within U2 snRNA. Both UV cross-linking experiments and mutational studies have provided considerable insight into the intricate network of interactions between pre-mRNA and snRNAs, and between the different snRNAs [13–15]. Upon spliceosomal assembly the extensive base-pairing between the U4 and U6 snRNAs is unwound, and U6 snRNA subsequently base-pairs with U2 snRNA and the 5´-splice site, thereby displacing U1 snRNA. The first trans-esterification reaction results in the cleavage of exon 1, and the formation of a lariat intron intermediate. A highly conserved loop in U5 snRNA aligns the 5´ and 3´ splice site for the second trans-esterification reaction [13,14,16]. Thus nuclear pre-mRNA splicing is a highly dynamic process involving transient RNA–RNA, protein–protein and RNA–protein interactions [13–20]. Spliceosome assembly is controlled by phosphorylation and dephosphorylation [21,22]. The structure of the assembled snRNAs and pre-mRNA within the active spliceosome bears some resemblance to that of group II self-splicing introns. This is consistent with the interesting idea that nuclear pre-mRNA splicing may be derived from group II intron self-splicing [1,2].
Spliceosomal proteins are divided into snRNP proteins that are tightly associated with snRNAs, and non-snRNP splicing factors (for reviews, see [6,9,10,25]). Protein components of the snRNPs are further classified into one of two groups: specific proteins that are found only in a given snRNP and the Sm proteins that associate with U1, U2, U4 and U5 snRNAs [6,10,25,26]. The Sm proteins assemble around a U-rich RNA sequence called the Sm site within U1, U2, U4 and U5 snRNAs and form a stable globular core domain in these snRNPs [27–29]. Some of the snRNP proteins contain previously identified sequence motifs, such as zinc fingers, helicase, protein kinase, GTPase and peptidyl/propyl cis–trans isomerases' motifs [6,10]." [s]
fast animation - spliceosome function : fast animation - spliceosome molecular action :
diagram - formation of a spliceosome : diagram - intron excision in mRNA precursors : diagram - pre-mRNA processing : animation ~ alternative splicing : animation of RNA splicing requires Flash Player plugin - Download plugin: clickable slide show - spliceosome intron removal : alternative splicing - click on fig 1 for animation : life cycle of an mRNA ~ click on Quicktime Q : clickable slide show - spliceosome intron removal :
Aberrant splicing creates mutant proteins, while alternative splicing generates complexity. Splice variants and epigenetic mechanisms account for the ability of about 30,000 human genes to code for about 100,000 human proteins. In addition to this variation, and that provided by recombination, each human gene possesses at least two isoforms – one from each parent. X Inactivation
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
No comments:
Post a Comment
Note: Only a member of this blog may post a comment.